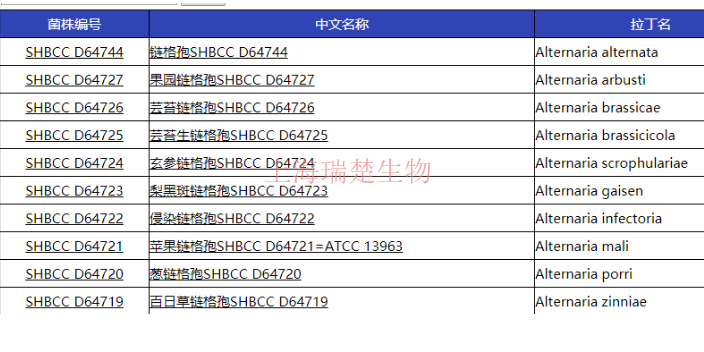
阿维菌素链霉菌菌株
枯草芽孢杆菌的芽孢在不同环境下的存活时间存在差异,主要受以下因素影响:1.**温度**:芽孢在高温下具有极强的耐受性。例如,芽孢能在120°C下存活20分钟,且能长期耐受60°C高温。然而,过高的温度会加速芽孢的失活。在适宜的温度下,芽孢可以保持活性数十年,甚至更长时间。2.**pH值**:芽孢在酸性或碱性环境中的耐受性不同。研究表明,pH值对芽孢的萌发和存活有影响。在酸性环境中,芽孢的萌发和存活能力降低。例如,pH值为4时,蜡样芽孢杆菌的芽孢萌发被完全抑制。3.**水分活度(Aw)**:水分活度对芽孢的存活和萌发也有重要影响。低水分活度条件下,芽孢的萌发被抑制,热抗性提高。例如,当Aw值降至0.92时,热与高压协同诱导的蜡样芽孢杆菌芽孢萌发与失活数量均降低几个数量级。4.**压力**:高压处理可以诱导芽孢萌发,降低其热抗性。例如,150MPa至600MPa的压力处理可诱导芽孢萌发,其中100~200MPa的压力处理通过引起芽孢内膜中的蛋白通道,诱导Ca2+-DPA释放,进而诱导芽孢萌发。
热红短芽胞杆菌(Brevibacillusthermoruber)在生物降解方面具有潜力,其具体作用机制和应用如下:1.**生物降解木质素**:-热红短芽胞杆菌在木质素降解方面表现出优异的性能。研究表明,该菌株能够在7天内降解81.97%的木质素,与木质素降解率相近。木质素的降解主要通过β-酮己二酸途径在37°C进行,而在55°C时,木质素的降解产物主要是苯甲酸物质,表明木质素是通过苯甲酸途径降解的。2.**高温耐受性**:-热红短芽胞杆菌能够适应高温环境,其营养体的生长温度在70℃以上,合适的生长温度为45-48°C。这种耐高温的特性使其在高温条件下的生物降解过程中具有优势。3.**代谢途径**:-热红短芽胞杆菌通过特定的代谢途径降解木质素。在37°C时,主要通过β-酮己二酸途径进行降解;而在55°C时,主要通过苯甲酸途径进行降解。这些途径的发现为木质素的生物降解提供了新的见解。4.**环境适应性**:-热红短芽胞杆菌在不同温度下的降解能力表明其在不同环境条件下的适应性。这种适应性使其在工业生产和环境修复中的应用具有潜力。5.**生物降解产物**:-热红短芽胞杆菌降解木质素产生的代谢产物,如苯甲酸,是堆肥中腐殖质形成的重要前体。 Pseudozyma antarctica菌种棉花黏液杆菌的细胞为革兰氏染色阴性、杆状,不产孢、不运动。

巴塞尔贪铜菌(Cupriavidusbasilensis)在生物修复重金属污染方面具有潜在的应用,尽管搜索结果中没有直接提及该菌种具体的生物修复机制。然而,基于其特性和β-变形菌纲的一些共性,我们可以推断其可能的生物修复机制:1.**重金属耐受性**:巴塞尔贪铜菌可能具有耐受多种重金属的能力,这使得它能够在重金属污染的环境中生存并发挥作用。2.**生物吸附**:该菌可能通过细胞表面的官能团与重金属离子形成稳定的复合物,从而吸收和固定重金属离子,减少其在环境中的迁移性和生物有效性。3.**生物转化**:巴塞尔贪铜菌可能具有将重金属转化为较低毒性形态的能力,例如将六价铬还原为三价铬,或将有机砷氧化为无机砷等。4.**植物-微生物联合修复**:巴塞尔贪铜菌可能与超积累植物形成共生关系,通过分泌植物生长调节物质和有机配位体,促进植物对重金属的吸收和转运,提高植物修复的效率。5.**分泌有机酸**:该菌可能通过分泌有机酸(如柠檬酸、琥珀酸等)与重金属离子络合,改变土壤中重金属的存在形态,降低其毒性并促进植物吸收。6.**产生铁载体**:巴塞尔贪铜菌可能通过产生铁载体与重金属离子络合,减少宿主植物对重金属的吸收,从而减轻重金属对植物的危害作用。
居中克吕沃尔氏菌(Kluyveraintermedia)在不同培养基上的生长特性可能表现出一些差异,但具体的信息在搜索结果中没有明确提及。然而,根据搜索结果,我们可以知道一些关于其在特定培养基上的特征:1.在**双倍乳糖胆盐培养基**中,居中克吕沃尔氏菌在45℃培养条件下不生长,这表明其对高温敏感。2.在**伊红美蓝琼脂培养基**上,该菌的菌落呈深紫黑色,圆形,边缘整齐,表面光滑湿润,并具有金属光泽。这些特征可以帮助实验室人员在进行菌株鉴定和分类学研究时,通过观察菌落的形态和颜色来识别居中克吕沃尔氏菌。尽管没有具体的不同培养基之间的比较数据,但上述信息提供了一些基本的生长特性,可以作为进一步研究的起点。如果需要更详细的不同培养基上的生长特性比较,可能需要通过实验室的实验来获得。橙色螺状菌通常生活在土壤中,尤其是草食动物的粪便、中性或微碱性的土壤、活树树皮以及腐烂的植物上 。
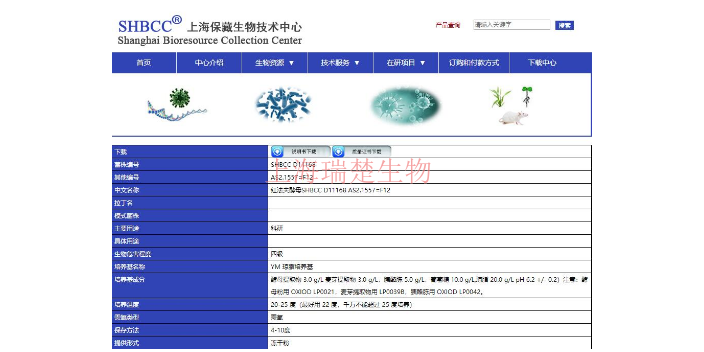
利用海考克氏菌的基因组信息来开发新的生物技术产品,主要可以通过以下几个步骤实现:1.**基因组测序与分析**:首先,需要对海考克氏菌的全基因组进行测序,以获得其完整的遗传信息。通过生物信息学工具分析基因组数据,识别潜在的生物合成基因簇(BGCs)和功能基因,这些可能与有用的生物活性物质的合成有关。2.**基因功能注释**:对预测的基因进行功能注释,确定它们在生物合成途径中可能的角色。这可以通过同源性搜索和比较基因组学来实现,以找到已知功能的相似基因。3.**基因编辑与敲除**:利用基因编辑技术(如CRISPR-Cas9系统)对特定的基因或基因簇进行敲除或修饰,以研究它们在生物合成过程中的作用,并可能激发沉默的生物合成途径。4.**异源表达系统**:将鉴定出的基因或基因簇在合适的宿主菌中进行异源表达,以产生目标化合物。这可能需要优化表达载体和培养条件,以获得高效表达和产物积累。5.**代谢工程**:通过代谢工程技术增强目标化合物的生物合成途径,可能包括增强前体供应、减弱副产物合成、或改变代谢流以提高产物的产率和质量。沉积物成对杆菌的丰富度和多样性与环境因子如盐度、亚硝酸盐和磷酸盐浓度明显有相关。铜绿假单胞菌PAO1菌种
沉积物成对杆菌能够适应不同的环境条件,包括在沉积物中存在有机质丰富的厌氧环境和含氧环境 。阿维菌素链霉菌菌株
拉氏根瘤菌(Rhizobiumleguminosarum)主要与豆科植物形成共生关系,进行固氮作用。其与非豆科植物共生的可能性相对较低,主要原因如下:1.**宿主专一性**:拉氏根瘤菌对豆科植物具有较高的宿主专一性,它们通过识别豆科植物释放的特定信号分子(如黄酮类化合物)来触发共生信号的交流。这种专一性使得它们很难与非豆科植物建立有效的共生关系。2.**信号交流障碍**:非豆科植物可能不产生或产生较少的根瘤菌所需的信号分子,导致根瘤菌无法识别并响应这些植物的信号,从而阻碍了共生关系的形成。3.**根瘤形成机制**:拉氏根瘤菌与豆科植物形成根瘤的机制是高度特化的。这种机制涉及到复杂的分子和细胞水平的相互作用,包括植物根部的细胞分化和根瘤菌的入侵。非豆科植物可能缺乏这种特化的机制,使得根瘤的形成变得困难。4.**固氮酶活性**:拉氏根瘤菌的固氮酶系统是针对豆科植物的共生固氮需求而优化的。这种系统在非豆科植物中可能无法有效发挥作用,因为不同植物对氮素的需求和利用方式可能存在差异。5.**生态位竞争**:在自然环境中,拉氏根瘤菌可能面临与其他微生物(如其他根瘤菌或非根瘤固氮菌)的竞争。阿维菌素链霉菌菌株